¿VIH o exosomas?

Imágenes tomadas al microscopio electrónico de exosomas aislados de la orina (izquierda) y de viriones de VIH aislado de un cultivo celular (derecha)
¿Por qué llamo a este artículo VIH o exosomas? Porque es una de las preguntas que hay que hacerse cuando se observan al microscopio electrónico células que están produciendo vesículas y/o virus. Existen similitudes entre los exosomas, y otras vesículas, y algunos virus. ¿Podemos diferenciarlos? Hagamos un repaso a la literatura científica para contestar esa pregunta.
¿Qué son los exosomas?
Los exosomas son vesículas que miden entre 30 y 90 nm de diámetro que son secretadas por un amplio rango de células eucariotas (Figura 1). El contenido de los exosomas está formado por proteínas y moléculas de RNA y su función es la de modular la función celular, por ejemplo presentando antígenos a las células T. A su vez contribuyen a la dispersión de enfermedades infecciosas y a la supervivencia de patógenos (Li y col, 2012) ya que la vía de producción de exosomas puede ser utilizada por diversos virus para gemar (salida de la célula arrastrando parte de la membrana celular de la propia célula) o bacterias que lanzan así señales moleculares a las células vecinas. Algunos ejemplos de virus que emplean esta vía son los retrovirus, como los virus que producen inmunodeficiencias en humanos, primates o felinos (Pelchen-Matthews y col., 2004), picornavirus, que produce la hepatitis A (Feng y col., 2013) o el virus de Epstein-Barr que produce la mononucleosis (Raab-Taub, 2012), entre otros.

Fig. 1A: Representación esquemática de las vesículas extracelulares. La proporción mayor corresponde a exosomas, microvesículas y restos de células que están muriendo (cuerpos apoptóticos). Es una simplificación, no todas las células producen todas estas vesículas. Fig. 1B: Tamaño comparado de las principales vesículas. Mientras que el tamaño de los exosomas es similar al de los virus, las vesículas coinciden en tamaño con algunas bacterias y con agregados proteicos. Los cuerpos apoptóticos alcanzan el tamaño de las plaquetas. Fuente: György y col. 2011
Cuando se realizan observaciones al microscopio electrónico de células crecidas in vitro en cultivo celular, se observan gran diversidad de vesículas de diversos tamaños (György y col. 2011). Un trabajo del grupo de Groopman llama la atención sobre este hecho (O’Hara y col., 1988). El grupo de Groopman lleva trabajando en el tema VIH/SIDA desde el descubrimiento de la enfermedad. En la época en la que todavía no se acaba de entender el mecanismo del SIDA, y la implicación del VIH el mismo, contribuyó a describir los efectos del virus sobre sus portadores (Groopman, 1985), el papel de las transfusiones de sangre como vía de transmisión del VIH (Groopman y col., 1984) o aislar el virus de presuntos enfermos de SIDA (Ho y col., 1985). Desde entonces ha publicado casi 200 trabajos, relacionados con la búsqueda de tratamiento y vacuna contra el SIDA, así como el análisis de la relación del VIH con la enfermedad. En un trabajo publicado en la revista “Human Pathology” en 1988 señalaron que en muestras procedentes de enfermos que poseían inflamación de ganglios, pero parecía no relacionado con el VIH, se observaban morfologías similares al VIH. En concreto analizaron al microscopio electrónico 20 muestras procedentes de ganglios de portadores del VIH y Encontraron partículas víricas en 18 casos. Todas ellas reaccionaron de forma positiva a anticuerpos anti-P24, una proteína del VIH. También observaron 13 partículas con morfología similar al VIH en 15 muestras de ganglios procedentes de personas no infectadas, pero ninguna de ellas reaccionó frente al anticuerpo anti-P24. La conclusión que obtienen es que sólo se observaron partículas compatibles con el VIH en ganglios de personas infectadas, pero deja de manifiesto que los viriones poseen morfología similar a vesículas que producen las células sanas. Esto es al menos lo que indica el resumen del artículo, desgraciadamente no he podido acceder al contenido completo del mismo, ya que no está disponible de forma gratuita. De todas formas en la red sí es posible encontrar la siguiente imagen de dicha publicación:

Figura 2: Fotografía obtenida al microscopio electrónico de células obtenidas de ganglios procedentes de un paciente portador del VIH (VIH +) y de un no portador (VIH -). Fuente: O’Hara y col., 1988
Lástima no poder acceder al artículo completo porque estas imágenes son poco informativas. Si bien es cierto que en el recuadro amplificado aparecen dos figuras muy semejantes, en la imagen completa (no ampliada) no puedo localizar este tipo de imagen. Ni siquiera queda muy claro cuál es el contorno celular debido al enorme número de vesículas flotantes. Tampoco se observan gemaciones, sea por parte de virus o de exosomas. Pero en cualquier caso da una información a tener muy en cuenta: se pueden encontrar vesículas de morfología similar a un retrovirus en muestras de individuos que posiblemente no sean portadores.
Un trabajo posterior mostró que la purificación de los viriones mediante ultracentrifugación en gradiente de sacarosa no parece mejorar el asunto, ya que las fracciones de aislamiento que debían contener el virus aparecen contaminadas con un alto número de vesículas y exosomas (Gluschankof y col., 1997).
En resumen, los exosomas y el VIH poseen un aspecto similar en preparaciones al microscopio electrónico y ambos se pueden encontrar en los métodos convencionales de purificación de retrovirus (ultracentrifugación en gradiente de sacarosa). Entonces, ¿el VIH y los exosoma son la misma cosa?, ¿no hay manera de distinguirlos?, ¿lo que observamos en células infectadas y no infectadas es exactamente lo mismo? Para contestar a estas preguntas debemos recurrir a los trabajos que estudian la morfología y composición bioquímica de exosomas y del VIH, tanto en células infectadas como no infectadas, así como a las técnicas de purificación y separación de exosomas y viriones. Vamos a ello.
Un buen punto de partida lo encontraríamos en la frase contenida en la Introducción de una publicación del grupo de S.J. Gould (el creador del concepto exosoma como caballo de Troya) que dice:
El virus VIH forma partículas en la misma zona que los exosomas, tiene la misma topología que los exosomas, tiene el mismo tamaño que los exosomas y están enriquecidos con las mismas moléculas que los exosomas.
.
Gould, de esta forma enfatiza que el VIH se comporta como los exosomas, no sólo eso, sino que según su hipótesis, los emplea como el vehículo para pasar de célula a célula, valiéndose de ellos cómo protección contra la respuesta inmune del huésped (Cantin y col., 2008). Pero este énfasis es exagerado, ya que hay suficientes datos en la literatura científica para afirmar que hay diferencias morfológicas y bioquímicas para distinguir exosomas y viriones. Repasemos algunas de esas diferencias:
Separación de exosomas y viriones del VIH
Como ya he comentado, tanto exosomas como viriones se co-localizan en la misma banda del gradiente de sacarosa formado tras una ultracentrifugación. Es por ello que diversos grupos de investigación se aprestaron a conseguir métodos de separación diferentes. El grupo de Caroline Gilbert, ha establecido un protocolo que permite separar físicamente los viriones de VIH de los exosomas, en células que han sido infectadas (Cantin y col., 2008). En este trabajo se informa que los exosomas comparten algunas características físicas y bioquímicas con los retrovirus debido que éstos emplean la maquinaria de producción de dichas vesículas en el proceso de gemación. Muestran que el tamaño de los exosomas es bastante variable, de 30 a 100 nm, mientras que los viriones de VIH poseen unos 100 nm de media. Vuelven a recordar que mediante centrifugación en gradiente de sacarosa es muy complicado realizar una separación completa, ya que los exosomas sedimentan entre densidades de 1.13-1.21 g/l, mientras que las partículas de VIH lo hacen a 1.16-1.18 g/l. Por tanto las preparaciones de VIH obtenidas por ese método estarían acompañadas de exosomas. Pero la aportación del equipo de Cantin es la separación diferencial en gradiente de Optiprep. Variando la concentración de este compuesto se pueden separar exosomas de viriones tal y como indica la figura:
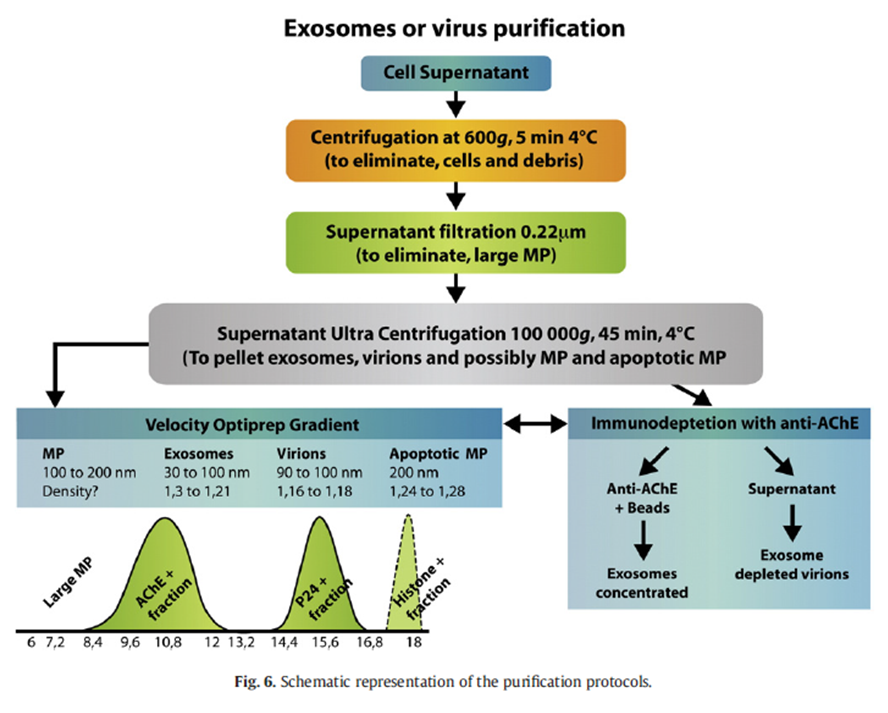
FIGURA 3: Protocolo de purificación de viriones de VIH. Fuente: Cartin y col., 2008
Como método de discriminación de ambas partículas (exosoma y VIH) se siguieron una serie de controles tras la purificación: (1) la acetilcolinesterasa, una enzima que se expresa, entre otras células, en eritrocitos y linfocitos. Esta enzima se ha localizado en exosomas con anterioridad. Este marcador sólo aparece en la fracción de concentración de Optiprep de 8.4-12, (2) la fracción de 14.4-16.8 contiene los viriones, los cuales no poseen actividad acetilcolinestarasa, es una fracción infectiva y posee morfología homogénea al microscopio electrónico. También es muy interesante hacer notar dos informaciones presentes en este trabajo: (1) se pueden usar anticuerpos anti-acetilcolinesterasa para inmunoprecipitar los exosomas, produciéndose así una fracción de viriones de alta pureza y (2) se emplearon dos grupos de linfocitos T, uno de ellos fue infectado con VIH y otro no (control). Solamente se detectó las proteínas p24 en las fracciones de purificación procedente en las muestras tomadas de células infectadas.
Otro trabajo describe el uso de anticuerpos frente a la proteína CD45, la cual es un antígeno común de la membrana de los leucocitos, abundante en las microvesículas, pero que no aparece incorporado en los viriones. Empleando anticuerpos contra CD45 (suministrados en micropartículas) se pueden inmunoprecipitar de forma mayoritaria las vesículas, de preparaciones de células infectadas con el VIH. De esa forma se obtienen una preparación enriquecida en VIH, donde las microvesículas prácticamente han desaparecido (Figura 4; Trubey, C.M. y col., 2003)

Figura 4: Purificación de viriones de VIH mediante inmunoprecipitación con anticuerpos frente a la proteína CD45, presente en exosomas. A la izquierda fracción obtenida por ultracentrifugación en gradiente de sacarosa antes del tratamiento y a la derecha parte no precipitada tras el tratamiento con anti-CD45. La barra de la escala representa 200 nm. Puede observarse que tras el tratamiento aparece una muestra de tamaño y morfología homogénea. Fuente: Trubey, C.M. y col., 2003
Otra técnica de separación que ha sido empleada es la cromatografía en columnas de Sepharosa CL-4B. Mediante esta técnica las microvesículas son retenidas, eluyen a menor velocidad que los viriones (Fuchigami, T., y col. 2002). En resumen se han desarrollado una serie de técnicas que permiten diferenciar las vesículas y exosomas de los viriones.
Diferencias morfológicas
Como ya he indicado, algunas publicaciones hablan sobre las similitudes morfológicas entre exosomas y viriones de VIH. Sin embargo un análisis del registro bibliográfico muestra que sí se observan algunas diferencias bien marcadas: (1) los viriones maduros poseen tamaño más uniforme que los exosomas, (2) se observan espículas alrededor de los viriones, (3) incluyen una cápside cónica en el 93% de las ocasiones y cilíndrica en el 7% de las mismas. Tendrían un aspecto como el de la siguiente Figura 5:

Figura 5. Varias partículas del VIH. Fuente: Servicio de microscopía electrónica de la Facultad de Biología de la Universidad de Leeds
Es cierto que estas morfologías son mucho más difíciles de apreciar (aunque se pueden observar) cuando se analizan células gemando, ya que esas muestras poseen un elevada proporción de vesículas y exosomas, así como partículas víricas inmaduras, pero sí que son perfectamente apreciables en muestras donde se han realizado un paso extra de purificación (como en la figura 5). Contamos en la actualidad con publicaciones que muestran la morfología y composición del contenido de los viriones, así como también se ha seguido la morfología del ciclo del virus, analizada incluso con la forma que poseen diferentes mutantes en proteínas clave del virus. También se han aplicado técnicas de crio-microscopia electrónica que exige la vitrificación de la muestra. Esta técnica produce cortes a diferentes profundidades y la reconstrucción de la imagen tridimensional mediante solapamiento de todas las imágenes obtenidas.
Por otra parte no he sido capaz de encontrar ningún trabajo donde se muestren exosomas purificados procedentes de células no infectadas con el VIH u otro virus (o no transfectadas con genes que expresen alguna de las proteínas víricas), que posean en su interior estructuras cónicas o espículas alrededor de su contorno como las de la figura anteriormente mostrada.
Diferencias bioquímicas
Para buscar si se han identificado diferencias bioquímicas entre los exosomas y los viriones he seguido dos estrategias: (1) Comprobar si existe alguna publicación que compare las vesículas/exosomas formadas por células infectadas y por células no infectadas con el VIH. Si existe algún trabajo de ese tipo habrá que ver si la comparación se hace a nivel bioquímico y (2) comprobar si hay publicaciones donde trabajando con células infectadas con el VIH (o que hayan sido tranfectadas para expresar alguna proteína del VIH) separan físicamente los viriones de los exosomas y analizan su composición química.
Una publicación del año 2012 entraría dentro del requisito de búsqueda #1. En ella se analiza mediante proteómica las vesículas de linfocitos (línea celular C9) que en unos casos fueron infectados con la variedad VIHNL4.3 del virus VIH durante 7 días, mientras que otros no fueron infectadas con dicho virus (muestra control). La infección se monitorizó mediante la cuantificación de la proteína p24 en el sobrenadante de las células infectadas. Este trabajo determinó diversas proteínas que eran producidas de forma diferencial en las células infectadas con el VIH. Los autores pudieron identificar las proteínas p24 y Tat del VIH en los exosomas producidos por las células infectadas (Liu y col., 2012), pero no en células no infectadas. Este resultado apoya que las proteínas del VIH sólo se expresan en células infectadas con el virus.
El grupo de S.J. Gould ha aportado algunos datos que encajaría dentro de los requisitos de búsqueda del punto #2. Este grupo estableció, como ya he comentado, la hipótesis del caballo de Troya. Según la misma, el VIH emplea la ruta de tráfico de los exosomas para pasar de célula a célula o bien se apoya en ella para gemar fuera de la célula. En una extensa (por el número de experimentos) publicación intentan establecer las proteínas que forman parte de los exosomas. Para ello tras el crecimiento de las células T se aislaron los exosomas formados y se analizó la abundancia de algunas proteínas. Así por ejemplo se identificaron las proteínas CD81 y CD63 en las membranas de los exosomas. Posteriormente siguieron el mismo protocolo estudiaron los exosomas producidas por células T transfectadas, que producían la proteína Gag-GFP, una variante de la proteína Gag del virus VIH fusionada a la proteína fluorescente GFP, para poder monitorizar mediante microscopía de fluorescencia el movimiento de la proteína en la célula que la produce. Pudieron comprobar que la proteína Gag-GFP expresada en células T se detectaba en las membranas de los exosomas, llegando a ser secretada de la célula por esta vía. Este resultado apoyaría la hipótesis del caballo de Troya (Booth y col., 2006; Fang y col., 2007).
La idea de que se pueden encontrar material del VIH en exosomas de células infectadas se vería reforzada por otro trabajo que sugiere que la proteína Nef del VIH, que está implicada en apoptosis (muerte) de la célula, y es una proteína clave en tomar el control de la célula infectada, es exportada por los exosomas y puede pasar de célula a célula (Lenassi , M. y col. (2010).
Incluso un reciente trabajo, que emplea la centrifugación en gradientes de OptiPrep (conocido también como iodixanol), para separar los exosomas de los viriones de VIH, muestra que las fracción de exosomas contiene RNA-VIH que no ha sufrido splicing, pero en los exosomas no se ha identificado RNA-VIH procesado (Cabezas y Federico, 2013). El hecho de que los exosomas sean capaces de contener RNA está permitiendo desarrollar técnicas biotecnológicas en las que se emplean los exosomas en terapia, usándolos como nanopartículas portadoras de ARN de interferencia (György y col., 2011). El ARN de interferencia puede paralizar (dependiendo del tipo) procesos celulares, impidiendo la transcripción (núcleo), o destruyendo el ARN mensajero que va a ser traducido (citoplasma), lo que puede ser de gran ayuda en terapia génica o para combatir infecciones víricas.
Conclusiones
Pienso que los datos aportados aquí, que a su vez proceden de publicaciones científicas que se puede encontrar mediante buscadores tipo PubMed, evidencia la existencia de dos entidades diferentes: los exosomas y el virus VIH. Los exosomas son microvesículas que forman algunas células sometidas a situación de estrés, mientras que el VIH es un retrovirus cuya biología molecular es bien conocida. Si bien los exosomas y el VIH pueden llegar a ser confundidos morfológicamente y comparten características bioquímicas, ambos se pueden separar mediante protocolos de purificación adecuados, lo que ha permitido entender con mayor profundidad que tienen en común (a nivel bioquímico) y cuáles son sus diferencias. Para finalizar, y a modo de anécdota, decir que me ha sorprendido mucho encontrar que algunas de las publicaciones que aquí presento eran usadas para apoyar tesis negacionistas sobre el VIH/SIDA, pero bueno ese es un tema en el que no voy a entrar, tal y como indiqué en un artículo anterior.
Referencias
Booth., A.M. y col. (2006) Exosomes and HIV Gag bud from endosome-like domains of the T cell plasma membrane J. Cell. Biol.
172:923-933.
Cabezas, S. y Federico, M. (2013) Sequences within RNA coding for HIV-1 Gag p17 are efficiently targeted to exosomes Cell Microbiol. 15:412-429.
Cantin, R. y col. (2008) Discrimination between exosomes and HIV-1: Purification of both vesicles from cell-free supernatants J. Immunol. Methods 338: 21-30.
Fang, Y. y col. (2007) Higher-Order Oligomerization Targets Plasma Membrane Proteins and HIV Gag to Exosomes PLoS Biol. 5: e158. doi:10.1371/journal.pbio.0050158.
Feng, Z. y col. (2013) A pathogenic picornavirus acquires an envelope by hijacking cellular membranes Nature. 496:367-371.
Fuchigami, T. y col. (2002) Acid-labile formylation of amino terminal proline of human immunodeficiency virus type 1 p24gag was found by proteomics using two-dimensional gel electrophoresis and matrix-assisted laser desorption/ionization-time-of-flight mass spectrometry Biochem. Biophys. Res. Comm. 293: 1107-1113
Gluschankof, P. y col. (1997) Cell Membrane Vesicles Are a Major Contaminant of Gradient-Enriched Human Immunodeficiency Virus Type-1 Preparations Virology 230:125-133.
György, B. y col. (2011) Membrane vesicles, current state-of-the-art: emerging role of extracellular vesicles Cell. Mol. Life Sci. 68: 2667-2688.
Groopman, J.E. (1985) Clinical spectrum of HTLV-III in humans Cancer Res. 45(9 Suppl):4649s-4651s.
Ho, D.D. (1985) Isolation of HTLV-III from cerebrospinal fluid and neural tissues of patients with neurologic syndromes related to the acquired immunodeficiency syndrome N. Engl. J. Med. 313:1493-1497.
Kadiu, I. y col. (2012) Biochemical and biologic characterization of exosomes and microvesicles as facilitators of HIV-1 infection in macrophages J. Immunol. 189:744-754.
Lenassi, M. y col. (2010) HIV Nef is secreted in exosomes and triggers apoptosis in bystander CD4+ T cells Traffic 11:110-122.
Li, M. y col. (2012) Quantitative proteomic analysis of exosomes from HIV-1-infected lympocytic cells Proteomics 12:2203-2211.
Groopman, J.E. y col. (1984) Virologic studies in a case of transfusion-associated AIDS N. Engl. J. Med. 31122:1419-1422.
O’Hara, C.J. y col. (1988) The ultrastructural and immunohistochemical demonstration of viral particles in lymph nodes from human immunodeficiency virus-related and non-human immunodeficiency virus-related lymphadenopathy syndromes Human Pathology 19: 545-549
Pelchen-Matthews, A. y col. (2004) Endosomes, exosomes and Trojan viruses Trends Microbiol. 127:310-316.
Raab-Taub, N. (2012) Novel mechanisms of EBV-induced oncogenesis Curr. Opin. Virol. 24:453-458.
Trubey, C.M. y col. (2003) Quantitation of HLA Class II Protein Incorporated into Human Immunodeficiency Type 1 Virions Purified by Anti-CD45 Immunoaffinity Depletion of Microvesicles J. Virol. 77:12699-12709.
Willey, R.D. y Gummuluru (2006) Immature dendritic cell-derived exosomes can mediate HIV-1 trans infection Proc. Natl. Acad. Sci. USA 103:738-743.

Últimos comentarios
| Eduardo Baldu en En EEUU los hospitales privado… | |
| Anónimo en Si la ciudadanía no comprende… | |
| Eduardo Baldu en Si la ciudadanía no comprende… | |
| m64bhkhfyj en Si la ciudadanía no comprende… | |
| Anónimo en La desforestación selectiva pu… | |
| Eduardo Baldu en ¿Los humanos somos el centro d… | |
| ateo666666 en ¿Los humanos somos el centro d… | |
| Anónimo en ¿Los humanos somos el centro d… | |
| ateo666666 en El peligroso engaño de que la… | |
| Anónimo en El peligroso engaño de que la… | |
| Eduardo Baldu en ¿Los humanos somos el centro d… | |
| Anónimo González en ¿Los humanos somos el centro d… | |
| ateo666666 en ¿Los humanos somos el centro d… | |
| luisgonzalesm en ¿Los humanos somos el centro d… | |
| ateo666666 en ¿Los humanos somos el centro d… |

Todos los textos de La Ciencia y sus Demonios se ofrecen bajo licencia Creative Commons Reconocimiento-NoComercial-SinObraDerivada 3.0 .



ClustrMaps

En línea ahora:
 "La Ciencia y sus Demonios" es miembro de la Red Escéptica
"La Ciencia y sus Demonios" es miembro de la Red Escéptica






Síguenos pulsando aquí

“Lo que puede ser afirmado sin pruebas, puede ser descartado sin pruebas”

Magnífico artículo, Manuel.
La cantidad de información que contiene es increible, de hecho creo que lo leeré alguna vez más para quedarme con todo lo posible, y además muy bien explicado.
Enhorabuena, por el trabajo.
Me gustaMe gusta
Muy buena entrada, como suele ser habitual, Manuel. Sólo un apunte rápido, creo que el hipervínculo de tu última frase » tal y como indiqué en un artículo anterior.» es incorrecto.
Me gustaMe gusta
Muy bueno, nos acerca mucho la metodología de trabajo y las complicaciones con las que os encontrais. Respecto a la hipótesis del caballo de troya, de ser cierta, me parece impresionante el grado de adaptación del virus (Espectacular la figura 5). Gracias por el artículo.
P.D: Sin ánimo de ser quisquilloso, párrafo anterior a la figura 2: «Encuentraron»
Me gustaMe gusta
Gracias por los comentarios y por indicar las erratas que habéis encontrado. Corregidas 😉
Me gustaMe gusta
Como dice el Papa de los negacionistas Peter Duesber en la revista negacionista Continuum del año 96: «HIV exists and has been isolated»
Un fragmento del artículo:
The existence of the retrovirus HIV predicts that HIV DNA can be
isolated from the chromosomal DNA of infected cells. This pre-
diction has been confirmed as follows: Full-length HIV-1 and HIV-
2 DNAs have been prepared from virus-infected cells and cloned
in bacterial plasmids13-15. Such clones are totally free of all viral
and cellular proteins, and cellular contaminants that co-purify
with virus.
Los estudios científicos citados (13-15):
13. Fisher AG, Collalti E, Ratner L, Gallo RC and Wong-Staal F: A molecular clone of
HTLV-III with biological activity. Nature (London) 316:262-265 (1985).
15. Barnett SW, Quiroga M, Werner A, Dina D and Levy JA: Distinguishing features of
an infectious molecular clone of the highly divergent and noncytopathic human
immunodeficiency virus type 2 UC1 strain. J. Virol. 67:1006-1014 (1993).
El artículo completo:
Haz clic para acceder a viewer
Me gustaMe gusta
http://www.ncbi.nlm.nih.gov/pubmed/1930645
«Antigen p24 was localized to HLA-DR positive cells that morphologically resembled macrophages in areas of villitis.»
Por favor, ruego se me clarifique esto. Gracias.
Me gustaMe gusta
Este es un fenómeno que no se ha acabado de entender, aunque algunas cosas se saben
(1) Las placentas (algunas células de ellas) expresan retrovirus endógenos (ERV), y algunas de esas proteínas son similares a las del VIH, de ahí que los anticuerpos contra algunas de esas proteínas den positivo. Eso no significa que mujeres embarazadas tengan anticuerpos contra todas las proteínas del VIH, mucho menos que esos anticuerpos estén circulantes en sangre.
(2) Esto no significa necesariamente que las embarazadas tengan anticuerpos contra esos ERV en sangre, normalmente no generamos anticuerpos contra nuestras propias proteínas (digo normalmente, porque las enfermedades autoinmunes consiste en generar anticuerpos contra nuestras propias estructuras).
(3) Las pruebas inmunológicas tienen deficiencias. Existen falsos positivos y falsos negativos. Eso pasa con la prueba del SIDA, con la de la tuberculosis (TB) o la de la hepatitis C. Hay casos de sífilis declaradas con negativos en las pruebas inmunológicas TB positivos sin rastro de tuberculosis. Es por ello que se requiere un segundo (al menos) criterio diagnóstico.
Asdf, me llama la atención que nunca comentéis los artículos sobre lanzáis preguntas. ¿Qué opinas del artículo bajo el que comentas, qué debilidad le encuentras?
Y ya que parece que dudas del VIH, me gustaría que me dijeras por qué si el virus no existe, y por tanto no hay nada que infecte células, ¿cómo se lleva a cabo terapia génica con el VIH? El virus se modifica genéticamente para poder restituir genes mutados en personas portadoras de dicha copia defectiva: http://lacienciaysusdemonios.com/2011/10/23/el-vih-como-herramienta-de-terapia-genica/
¿Cómo se hace eso con un virus que no existe? Por favor, clarifícame eso. Gracias
Me gustaMe gusta