Evolución en Euglena & company: de la depredación a la fotosíntesis.

Euglena viridis. Crédito: microagua (Flickr)
Un microbio típico de muchos libros de biología es Euglena. A veces es llamada alga porque en presencia de sales inorgánicas y una fuente de luz, es capaz de realizar la fotosíntesis como cualquier planta o alga verde. A veces es llamada protozoo, porque con materia orgánica y en oscuridad puede nutrirse como hacen los hongos y los animales sencillos. Es una entidad formada por una única célula, de color verde gracias a que tiene útiles cloroplastos de brillante color. Sin embargo, sus ancestros cercanos son fieros depredadores, sin cloroplastos e incapaces de realizar la fotosíntesis. Entonces… ¿Cómo Euglena se convirtió en lo qué es? A esta y otras cuestiones responderemos en esta entrada.
.
Los Euglenida, a los que también podemos llamar euglénidos, son un enorme grupo de microorganismos formados por una única célula que actualmente cuentan con más de mil especies descritas. Son habitantes de los diminutos espacios que quedan entre los granos de arena u otros sedimentos de los fondos de mares, ríos y lagos; aunque hay otros que prefieren vivir nadando en la columna de agua.
El nombre de euglénidos deriva de los términos griegos «Eu, verdadero» y «glēnē, ojo». Es un nombre extraño, concedido por el naturalista, zoólogo, botánico, anatomista, geólogo y microscopista alemán Christian Gottfried Ehrenberg (1795 –1876). Prolífico como pocos, seguramente se sentiría atraído por la mancha ocular que poseen estos organismos y por ello, la criatura que conocemos como Euglena recibía en 1830 su nombre, «ojo verdadero», por vez primera. Luego, dicho nombre sería extendido para denominar a todos sus parientes.
¿Qué es dicha mancha ocular? Aparte de ser un carácter ideal para su identificación, se trata de un sencillo tipo de ojo acorde con la idea de “ojo prototípico” de Darwin. Es tan simple que si un euglénido es en sí mismo una única célula, dicho “ojo” es un componente aún más simple que una célula. Aún así podemos decir que es una estructura sofisticada compuesta por una membrana que envuelve unas cuantas esferas formadas por pigmentos que absorben la luz. Al lado de esa bolsa llena de esferas se sitúa una estructura llamada fotorreceptor, de modo que si el fotorreceptor recibe luz solar filtrada a través de la mancha ocular, este fotorreceptor se «activa« y “dirige” el movimiento celular.
Para permitir tal movimiento los euglénidos tienen acoplados, cerca del fotorreceptor, una cosa llamada flagelos. Los flagelos son construcciones celulares que recordarían a enormes látigos, que al ser sacudidos impulsan al microbio en el medio líquido en el que vive. Los espermatozoides del primate pelado conocido como Homo sapiens sapiens tienen un solitario y único flagelo, que nosotros vemos como esa cola tan larga y móvil. Los euglénidos, por su parte, suelen tener dos flagelos, muchas veces orientados uno hacia delante y otro hacia atrás. Estos flagelos salen desde una especie de saco interno de la célula conectado con el exterior llamado «reservorio». También hay euglénidos como nuestra querida Euglena, que aunque tienen dos flagelos, solo uno es útil para impulsarse ya que el otro es de carácter vestigial.
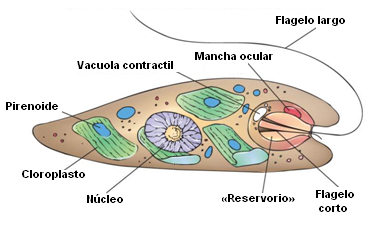
Esquema de la estructura básica de Euglena. Crédito: San Diego Mesa College
.
Criaturas de película.
.
Los euglénidos también han patentado una estructura única y muy compleja de gran interés para la microbiología. Dicha estructura en inglés recibe el nombre de «pellicle» y aunque no he localizado una traducción segura al castellano, en principio creo que un término apropiado debería de ser película. Hay dos razones por las que es interesante hablar de la película de los euglénidos: la primera, porque es una estructura que es diferente según el tipo de alimentación del microorganismo; la segunda, es que resulta precioso verla desde un prisma evolutivo.
Dicha película es visible en la superficie de la célula como una serie de estrías o canales muy marcados, estos arrancan en un extremo de la célula y a modo de líneas paralelas recorren toda la célula a lo largo de su longitud. Estos canales o estrías pueden seguir una línea recta o pueden seguir un recorrido espiral. Hay especies en las que tales canales o estrías acaban en un orificio que se introduce en el interior celular. Tales estrías también pueden estar tachonadas de poros que comunican con el interior. Y en algunas especies se sigue un patrón en el que el número de estrías disminuye conforme se acercan al principio y/o final de la célula ¿Qué función tienen? Posiblemente la película cumple un papel en la movilidad de los euglénidos, ya que no solo se mueven vía flagelos, sino que pueden modificar su cuerpo para reptar sobre las superficies. En algunas especies la película también desempeña un papel en la alimentación.

Detalle de la película de euglénidos. Izquierda, con microscopio óptico en Euglena, créd.: wiki. Centro, con microscopio electrónico de barrido, nótese las estrías de Lepocinclis ovum, créd.: BioEssays. Derecha, con microscopio electrónico de transmisión, loncha de Distigma proteus, nótese el canal (entre las dos flechas), créd.: Journal of Eukaryotic Microbiology.
.
A nivel íntimo ¿cómo está organizada dicha película? Su componente principal es una proteína que recibe el nombre de «articulina» («articulin» en inglés, para los que quieran profundizar). Esta proteína se organiza formando una serie de placas paralelas entre sí que recorren longitudinalmente la célula. Por su parte, estas placas encajan lateralmente unas con otras gracias a una serie de proyecciones; de modo que las proyecciones prearticulares de una placa encajan con las proyecciones postarticulares de la placa situada al lado. Entre una placa y la siguiente es donde se sitúa la estría o canal.
Encima de las placas de «articulina» tenemos a la membrana plasmática; en toda célula esta membrana es lo que separa el interior de la célula del exterior de la célula. Por su parte, debajo de las placas se sitúa un tubo formado por el retículo endoplasmático, este es un componente de la célula con múltiples funciones, desde transporte de variopintos productos hasta la fabricación de grasas, pasando por ser el último lugar por el que pasan las proteínas antes de estar terminadas. Finalmente, las inmediaciones de los canales o estrías están rodeadas de microtúbulos, una estructura que forma parte del “esqueleto” de la célula y que entre otras cosas, permite su cambio de forma.

Estructura básica de la película de los euglénidos. Crédito: BioEssays.
.
Las películas de los euglénidos, similares en su esquema básico, ofrecen una amplia diversidad de patrones y formas. Sin embargo, a pesar de su complejidad hay biólogos que se han atrevido a escudriñar su evolución a lo largo de los distintos linajes de euglénidos. Así nos encontramos con el Dr. Brian S. Leander, profesor de la University of British Columbia (USA), un especialista en la ecología y evolución de diversos protozoos unicelulares, como los euglénidos, los dinoflagelados y los cercozoos. De todos sus trabajos publicados en este ámbito destaca uno año 2007 en la revista científica BioEssays, titulado “Macroevolution of complex cytoskeletal systems in euglenids”, que resume los avances científicos más importantes en el estudio evolutivo de la película de los euglénidos y del que daremos cuenta en esta entrada.
¿Cuál fue el origen de la película de los euglénidos? Bueno, hasta ahora hay pocas respuestas. Los científicos hoy reconocen a dos grupos de microbios acuáticos como los parientes vivos más cercanos de los euglénidos: los kinetoplástidos (como Trypanosoma, el causante de la “enfermedad del sueño”) y los diplonémidos. Al igual que ellos, los primeros euglénidos no debieron de tener película y dicho elemento es invento suyo. Posiblemente, esta pudo formarse como una delgada capa de proteínas situada debajo de la membrana celular ¿Por qué desarrollar una proto-película? No se sabe, quizás permitió una mayor facilidad para reptar sobre el sustrato.
Esta “proto-película” tendría una “proto-estría”, el lugar donde se resguardaría uno de los flagelos. Ahora queda aumentar el número de estrías ¿Cómo hacerlo? Una posible forma de lograrlo es mediante divisiones celulares fallidas, ya que en estos seres el número de estrías se duplica antes de comenzar a dividirse. Así, este fenómeno pudo convertir al hipotético ancestro de una estría en un segundo e hipotético ancestro de dos estrías: el ancestro de los actuales euglénidos bacterívoros (cazadores de bacterias).
Una segunda duplicación del número de estrías originaría los más primitivos euglénidos bacterívoros de hoy día, que presentan en sus sencillos cuerpos únicamente 4 estrías. Estas duplicaciones volverían a pasar durante las primeras etapas evolutivas de los actuales euglénidos bacterívoros (que pasaron de 4 a 8 estrías); en la transición desde euglénidos bacterívoros a eucarívoros (saltando de 10 a 20 estrías), durante la propia evolución de los eucarívoros (de 20 a 40 estrías) y en la evolución interna de los fotosintéticos (donde algunos pasaron de 60 a 120 estrías).

Evolución en el número de estrías en la película de los euglénidos (P: número de estrías). Crédito: BioEssays.
.
¿Por qué aumentaron y modificaron tanto el número de estrías o canales de la película? Seguramente la respuesta estriba en el tipo de alimentación de cada euglénido y en la posibilidad de explotar distintos recursos de forma cada vez más eficiente. Así los euglénidos cazadores de bacterias (bacterívoros) son relativamente rígidos y cuentan con pocas estrías (de 4 a 12). Sus descendientes son los euglénidos depredadores de grandes células que no son bacterias (eucarívoros), estos requieren de cuerpos más flexibles para cazar a sus presas y engullirlas, por ello cuentan con una gran cantidad de estrías (de 20 a 56), ya que tales cifras permiten un cuerpo mucho mejor articulado. Desde los eucarívoros evolucionaron ciertos euglénidos osmótrofos (se limitan a absorber nutrientes), donde algunos como Distigma conservan la flexibilidad (con 18 a 22 estrías) y otros como Rhabdomonas y Menoidium han reducido el número de estrías (tienen de 8 a 14), siendo menos articulados y por lo tanto, más rígidos; lo cual no les supone problema ya que su modo de vida, limitado a absorber los nutrientes disueltos en el agua, no requiere de cuerpos flexibles.
También evolucionados desde euglénidos eucarívoros, esos depredadores de grandes células que no son bacterias, encontramos las especies fotosintéticas. Ellas presentan una gran diversidad de formas, incluyendo especies muy flexibles y otras muy rígidas. Entre las flexibles se encuentran las más primitivas Eutreptiales y otras como Euglena y Colacium, así como aquellas que se recubren a sí mismas de una cáscara protectora como Trachelomonas y Strombomonas; todas ellas tienen entre 40 y 50 estrías, aunque las hay con 120, como Euglena obstusa. Por su parte, las especies rígidas cuentan desde 15 hasta 20 estrías, incluyen seres que ya nombramos cuando hablamos de la evolución de la película, como Phacus, Lepocinclis y Monomorphina.

Algunos ejemplos de euglénidos. G: Peranema trichophorum (50 estrías). J: Euglena mutabilis (40 estrías). K: Euglena sp. (40 estrías). L: Monomorphina ovata (16 estrías). O: Phacus sp. (32 estrías). Crédito: Crédito: BioEssays.
.
Estos microbios también modificaron la propia estructura de la película. La más simple y menos rígida pertenece a varios euglénidos depredadores y otros que se alimentan absorbiendo nutrientes del medio. Esta película está formada por placas sin ningún tipo de proyección y limitadas a mantener algún que otro poro (A). En los euglénidos fotosintéticos más primitivos (ej. Eutreptiales) aparece un preámbulo de proyección prearticular en forma de filamentos delgados y escasos, asimismo aparece una proyección postarticular en forma de peine (B). El aumento de la robustez de dichas proyecciones aumenta la rigidez de la célula. Así en algunas especies de Euglena dichos filamentos se engrosan formando una serie de proyecciones lineares (C), que evolucionan a un aspecto similar a dientes en criaturas de cuerpos rígidos como los géneros fotosintéticos Phacus, Lepocinclis y Monomorphina (D). El siguiente paso es una proyección postarticular con forma de placa, cuya siguiente mejora es el reforzamiento con “costillas”; ambos tipos de proyección postarticular aparecen en varias especies de Lepocinclis (E y F). Tales modificaciones en la placa aumentan la estabilidad de la película y permiten que las células puedan aumentar su tamaño. Este proceso evolutivo solo se observa en los euglénidos fotosintéticos; si bien sus razones últimas aún son una incógnita.

Evolución de las placas de la película de los euglénidos (ver texto anterior). Crédito: BioEssays.
.
Sospechas confirmadas.
.
Todo esto está muy bien, pero ¿tales explicaciones tienen algún apoyo más aparte de la propia forma que adquiere la película o el número de estrías que podemos ver? Pues va a ser que sí. Ya que estos estudios no solo encajan muy bien con las características corporales de los euglénidos, sino además con su propia genealogía, conocida a través de los estudios genéticos realizados en este grupo.
Así en la imagen inferior contamos con dicha genealogía. Los distintos números representan las novedades evolutivas que han ido apareciendo: una característica de los euglénidos y sus parientes más próximos es la presencia de dos flagelos que parten de una invaginación de la célula llamada «reservorio», uno de los flagelos se dirige hacia atrás y el otro puede hacerlo hacia delante, introduciéndose en un surco de la célula llamado «sulcus» (1); los euglénidos por su parte han inventado la película que caracteriza a este grupo (2). La siguiente innovación aparece en euglénidos bacterívoros como Ploeotia y Entosiphon, estos poseen un sofisticado sistema de alimentación basado en “varillas” y “paletas” (del que hablaremos posteriormente) (3); dicho sistema sería heredado por los euglénidos eucarívoros como Dinema, de cuerpos mucho más flexibles para poder ingerir presas mucho más grandes (4). En el siguiente punto encontramos euglénidos eucarívoros altamente flexibles, como Peranema y Urceolus (5), de ellos se originan los primeros euglénidos fotosintéticos, los cuáles en su versión más primitiva desarrollan filamentos en las placas (de la película) que son la antesala de las proyecciones que vemos en otros grupos, tal y como puede hoy en las Eutreptiales (6). A partir de este momento tiene lugar una importante modificación de la película, sustituyéndose la movilidad reptante por una movilidad impulsada por el flagelo para explotar la columna de agua (7); y finalmente el desarrollo de una película más rígida y robusta (9) o la invención de una concha dura de carácter protector (8).

Filogenia (relaciones de parentesco) entre los distintos grupos de euglénidos (ver con texto anterior) realizada en base a los genes rRNA y «proteína del shock término 90». Crédito: BioEssays.
.
Notables cazadores de bacterias.
.
Para su alimentación los euglénidos cuentan con un elemento denominado aparato de alimentación, de las que existen distintas variantes y modelos, a cada cuál más complejo. En esta entrada lo hemos comentado un poco por encima, pero no estará de más repetirlo. Los euglénidos son criaturitas muy versátiles que han desarrollado cuatro tipos de alimentación: bacterívoros (comen bacterias y partículas de pequeño tamaño), “eucarívoros” (se comen microorganismos que no son bacterias y partículas de gran tamaño), osmótrofos (simplemente absorben nutrientes ya disueltos en el medio) y fotosintéticos (realizan la fotosíntesis como las plantas). Algunos, como la popular Euglena, son capaces de combinar más de un tipo de alimentación y son llamados «mixótrofos».
Como hemos podido ver en el apartado anterior, los más antiguos euglénidos con bacterívoros, depredadores de bacterias que ocasionalmente ingieren alguna partícula de reducido tamaño ¿De dónde salieron estos pequeños cazadores? Al parecer los parientes más próximos de los euglénidos son los kinetoplástidos, un grupo de microorganismos que incluye a peligrosos parásitos para el hombre, como Trypanosoma cruzi, presente en el continente americano y causante de la enfermedad de Chagas, cuyo vector es la chinche besucona o chinche picuda (Triatoma infestans); en el continente africano la preocupación se la lleva Trypanosoma brucei, causante de la enfermedad del Sueño y cuyo vector es la mosca tse-tse (Glossina sp.). Pero los Trypanosoma son kinetoplástidos altamente avanzados y especializados. La mayoría de los kinetoplástidos tienen otra forma de vida, muchos de ellos son discretos seres que reciben el nombre de bodónidos.
Los bodónidos se encuentran entre los kinetoplástidos más primitivos. Son un silencioso y amplio grupo de microorganismos donde la mayoría son microbios pequeñitos (en comparación con otros microbios) que viven felizmente cazando bacterias. Por su parte, los euglénidos primitivos (entre los que incluimos algunos como Petalomonas, Entosiphon y Ploeotia) también son microbios pequeñitos que viven felizmente cazando bacterias. Por ello los científicos imaginan que los ancestros de euglénidos y kinetoplástidos, ambos primos cercanos, debieron ser pequeños microbios cazadores de bacterias.

Kinetoplástidos. Izquierda, Bodo saltans, el cazador de bacterias de vida libre, créd.: EOL. Derecha, Trypanosoma brucei, parásito humano causante de la enfermedad del Sueño créd.: Nature
.
Para poder comerse a tales bacterias, estos microorganismos disponen del llamado aparato de alimentación tipo I. El más simple, presente en el euglénido Petalomonas y similar al de algunos bodónidos, consiste en una especie de sencillo saco conectado con el exterior llamado citofaringe. Dicha citofaringe está exteriormente reforzada con microtúbulos. Este saco se sitúa en posición ventral con respecto al «reservorio», ese punto del que salen los flagelos. Junto a la comunicación del «reservorio» con el exterior, se sitúa la propia apertura de la citofaringe con el exterior, a través de la cuál serán ingeridas las bacterias poco espabiladas.

Esquema del aparato de alimentación tipo I del cazador de bacterias Petalomonas. Crédito: TRENDS in Microbiology & BioEssays.
.
El aparato de alimentación tipo II es un poco más sofisticado y aparece en un euglénido bacterívoro llamado Ploetia. Sigue el mismo diseño que el anterior, solo que la citofaringe ahora aparece reforzada por una estructura proteica llamada “paletas” que rodea completamente a la citofaringe. También se incluyen dos varillas reforzadas por microtúbulos y protegidas por una matriz cementante. Estas varillas son extraordinariamente largas, extendiéndose a lo largo de toda la longitud de la célula.

Esquema del aparato de alimentación tipo II del cazador de bacterias Ploetia. Crédito: TRENDS in Microbiology & BioEssays.
.
El aparato de alimentación tipo III es una modificación del anterior y aparece en euglénidos como Diplonema ambulator o Dinema sulcatum. A lo largo de las distintas especies que tienen este sistema de alimentación puede observarse una sustitución gradual de la matriz cementante por un cada vez mayor número de microtúbulos, tal y como termina por observarse en Dinema. Al mismo tiempo, las “paletas” no rodean completamente la citofaringe.
Dinema también aparece como un caso transicional hacia una forma de vida eucarívora, ya que aunque depreda habitualmente sobre bacterias, también es capaz de engullir presas tan grandes como ella misma. Esta evolución hacia una dieta eucarívora también está asociada al incremento del número de estrías, ya que estas otorgan mayor número de puntos de articulación y por lo tanto, mayor flexibilidad, facilitando la capacidad para engullir presas grandes.
El más sofisticado es el aparato de alimentación tipo IV y aparece en un euglénido bacterívoro (sí, bacterívoro) llamado Entosiphon. Es una modificación del anterior, donde la matriz cementante ha desaparecido completamente (como sucede en algunos usuarios del aparato de alimentación tipo III) dejando paso a un reforzamiento basado en microtúbulos. Además, una de las varillas comienza a dividirse en dos desde del extremo posterior de la célula hasta el extremo anterior, por lo que da la apariencia de tener tres varillas.

Esquema del aparato de alimentación tipo III del cazador Dinema. Esquema del aparato de alimentación tipo IV del cazador Entosiphon. mv: microtúbulos de refuerzo de la varilla. Crédito: TRENDS in Microbiology & BioEssays.
.
Y cazadores de organismos eucariontes.
.
Como comentábamos anteriormente, Dinema es un representante de lo que podría ser un paso desde un organismo depredador de bacterias hacia uno cazador de organismos unicelulares eucarióticos o eucariotas. Es decir, aquellos seres formados por una única célula pero que no son bacterias; ya que aunque Dinema es depredador de bacterias, también es lo suficientemente flexible como para atrapar, envolver y engullir organismos eucarióticos, generalmente muchísimo más grandes que una bacteria.
El aparato de alimentación de los depredadores de microorganismos eucariotas es muy similar al aparato de alimentación de tipo III. Hemos visto que en los euglénidos este aparato presenta un amplio gradiente, desde algunos muy similares al tipo II, donde las varillas están cubiertas de una matriz cementante y están rodeadas por pocos microtúbulos; hasta el aparato de Dinema donde las varillas han perdido toda la matriz cementante y están rodeadas por un amplio conjunto de microtúbulos. Pues bien, algunos depredadores de eucariotas como Peranema aparece como un caso intermedio entre ambos, ya que ni tienen tan reducida la matriz cementante y al mismo tiempo han incrementado el número de microtúbulos que rodean a esta.
Otro rasgo a destacar es el hecho de que estos depredadores tienden a tener una película con un número de estrías muy elevado, mientras que los cazadores de bacterias son relativamente rígidos y no tienen mucho más de diez estrías. Por ello estos cazadores de eucariotas son muy flexibles y llegan a tener de veinte a cincuenta estrías. Es una característica muy importante ya que cada estría implica un punto de articulación, por lo que a mayor número de articulaciones mayor flexibilidad dispondrán sus cuerpos, facilitando la ingestión de presas. Asimismo, un mayor número de estrías permite que puedan ser depredadores mucho más grandes, lo que incrementa el rango de microbios sobre los que pueden depredar. Quizás debido a este incremento del tamaño, el aparato de alimentación no se extiende en toda la longitud de la célula, sino que solo se extiende un tercio de dicha longitud.

Esquema delaparato de alimentación tipo III del cazador de eucariontes Peranema. Crédito: TRENDS in Microbiology & BioEssays.
.
Convirtiendo un cazador en un ser fotosintético.
.
Los euglénidos fotosintéticos son descendientes directos de los euglénidos depredadores de eucariotas. Sin embargo, son muy pero que muy diferentes:
- Los depredadores tienen un aparato de alimentación bien desarrollado. Los fotosintéticos por su parte tienen un aparato de alimentación muy reducido y de carácter vestigial.
- Los fotosintéticos tienen una película que reduce su número de estrías conforme se aproxima al extremo posterior de sus cuerpos. Los depredadores no presentan este fenómeno, salvo el caso dudoso de Peranema.
- Los depredadores se desplazan arrastrándose por el sustrato mientras modifican sus cuerpos. Los fotosintéticos nadan en la columna de agua mientras sacuden sus flagelos.
- Los depredadores no tienen cloroplastos, carecen de cualquier tipo de maquinaria para realizar la fotosíntesis y tampoco poseen los pre-requisitos para realizarla. Los fotosintéticos por su parte tienen cloroplastos muy bien desarrollados y realizan la fotosíntesis la mar de bien.
Por lo tanto, la pregunta es evidente… ¿Cómo pudieron evolucionar los euglénidos fotosintéticos de los euglénidos depredadores si son tan claramente opuestos?

Comparativa de distintos euglénidos en función de la dieta. Están a escala. (1) Ploeotia vitrea, (2) Petalomonas cantuscygni, (3) Dinema sulcatum, (4) Peranema trichophorum, (5) Euglena geniculata, (6) Monomorphina ovata, (7) Phacus sp., (8) Lepocinclis sp. y (9) Lepocinclis oxyuris. Crédito: TRENDS in Microbiology.
.
Para muchos científicos, la respuesta se llama endosimbiosis. Se trata de un proceso en el que un organismo vive como simbionte en el interior del cuerpo de otro organismo llamado hospedador. Este simbionte, con el paso del tiempo, termina perdiendo su independencia, convirtiéndose en un elemento más del hospedador.
Dicho modelo recibe el nombre de Teoría Endosimbiótica; cuya fama se la debemos a la fallecida bióloga estadounidense Lynn Margulis (1938 – 2011) y no solo explica muy bien el origen de las mitocondrias y cloroplastos de las células eucariotas a partir de bacterias libres, sino que la cantidad de pruebas acumuladas a favor de dicho modelo es abrumadora. Por otro lado, este modelo suena perfectamente bien para explicar el gran “salto” desde un euglénido depredador hasta un euglénido fotosintético, al menos en lo que a la parte de la fotosíntesis se refiere. Pero… ¿cómo explicarlo en la práctica? ¿Se han encontrado euglénidos que alojen algas simbiontes en su interior?
En este año 2012 fue publicado en la revista científica BMC Evolutionary Biology un artículo dirigido por el equipo de investigación del Dr. Brian S. Leander que responde perfectamente a tales preguntas. En dicho trabajo se había estudiado un euglénido depredador recientemente descubierto bautizado como Rapaza viridis, un animal que tiene todos los puntos para ser una etapa transicional entre un modo de vida depredador y otro fotosintético.

Rapaza viridis merendándose un alga Tetraselmis. Izquierda, con microscopio óptico, alga señalada con la flecha. Nótese las estrías en el borde del cuerpo de Rapaza, escala de 5 μm. Derecha, con microscopio electrónico de barrido, alga señalada con la flecha, escala de 5 μm. Crédito: BMC Evolutionary Biology.
.
El primer rasgo que separa un poco a Rapaza de otros euglénidos depredadores es que este, al igual que los euglénidos fotosintéticos, podía desplazarse alegremente por la columna de agua gracias a sus flagelos. Porque Rapaza es un depredador y su dieta consiste en microalgas unicelulares, como las del género Tetraselmis, las cuáles tienen flagelos y nadan alegremente en la columna de agua. Aunque Rapaza puede desplazarse con sus flagelos en el agua, una vez captura su presa emplea el mismo movimiento reptante que los demás euglénidos depredadores para engullir su presa, en un proceso que dura de 5 a 40 minutos; cuando dicho proceso finaliza, Rapaza vuelve a moverse con sus flagelos. Los científicos también anotaron que este depredador no puede sobrevivir más de 35 días sin cazar una presa.
Pero Rapaza viridis es mucho más interesante, porque resulta que no es únicamente un depredador. Sino que conserva en su interior dos tipos de cloroplastos: los de las algas Tetraselmis que depreda y los propios de otros euglénidos fotosintéticos. Y son cloroplastos funcionales, porque los experimentos demuestran que sin luz no sobrevive más de una semana.
Así que tenemos un euglénido depredador que necesita cazar y que también necesita hacer la fotosíntesis para vivir. Además no solo ha aprendido a conservar los cloroplastos de las algas que captura, sino que también tiene sus propios cloroplastos. Y aunque usa los flagelos para nadar también sabe reptar como cualquier depredador… Por lo que es un ser situado justo en el punto medio de la conversión de un depredador a un ser fotosintético. Resulta tan interesante que realizar más preguntas es una inercia imposible de frenar.
¿Qué se dice de su aparato de alimentación? Qué está justo en el medio, ni tan desarrollado como un depredador nato ni tampoco tan vestigial y reducido como en un fotosintético de toda la vida… ¿Y de su película? Qué es el primer depredador que revela claramente un patrón de reducción de estrías hacia el extremo posterior de su cuerpo, exactamente tal y como se espera del ancestro de los más primitivos euglénidos fotosintéticos, los Eutreptiales…
Vamos, una criatura que ni regalada. Para terminar, ¿los estudios genéticos dirán algo interesante, verdad? Verdad. Confirmando toda sospecha, Rapaza viridis se sitúa justo antes de los Eutreptiales, los más primitivos fotosintéticos; y justo después de los depredadores de los que descienden dichos fotosintéticos.
Rapaza viridis como elemento transicional en la evolución de los euglénidos. Cl: cloroplasto; Pm: paramilo. Crédito: BMC Evolutionary Biology.
.
En conclusión, nos ha tocado la lotería. Y aquí lo dejamos. A lo largo de esta entrada hemos visto que los euglénidos, ese amplio conjunto de microorganismos, tienen una diversidad y disparidad de formas asombrosa. Espero que me sea perdonado que el titular de la entrada solo sea una pequeña parte de todo el artículo, si puedo excusarme, diría que los euglénidos tienen muchos rasgos que pueden estudiarse estupendamente desde un punto de vista evoutivo: aumento de la complejidad de la película, variación en el número de estrías y la modificación del aparato de alimentación; todo ello confirmado con la genética. Finalmente, hemos visto como un depredador estricto puede pasar a convertirse en un ser principalmente fotosintético. Una maravilla, sin embargo el estudio de Rapaza viridis aún está en pañales, por ello… ¡son necesarios más estudios!
.
Entradas relacionadas:
.
BONUS TRACK. Euglena viridis. Nótese la mancha ocular de color rojo.
.
Referencias.
- Barsanti y col. 2012. Fundamental questions and concepts about photoreception and the case of Euglena gracilis. Integrative Biology 2: 22-36.
- Deschamps P y col. 2011. Phylogenomic Analysis of Kinetoplastids Supports That Trypanosomatids Arose from within Bodonids. Molecular Biology and Evolution 28 (1): 53-58.
- Leander BS. 2004. Did trypanosomatid parasites have photosynthetic ancestors?. TRENDS in Microbiology 12 (6): 251–258. Artículo disponible completo también aquí.
- Leander BS & Farmer MA. 2000. Comparative Morphology of the Euglenid Pellicle. I. Patterns of Strips and Pores. Journal of Eukaryotic Microbiology 47 (5): 469-479. Artículo completo también disponible aquí.
- Leander BS & Farmer MA. 2001. Comparative Morphology of the Euglenid Pellicle. II. Diversity of Strip Substructure. Journal of Eukaryotic Microbiology 48 (2): 202-217. Artículo disponible también aquí.
- Leander BS y col. 2001. Character evolution in heterotrophic euglenids. European Journal of Protistology 37 (3): 337-356. Artículo completo también disponible aquí.
- Leander BS y col. 2001. Trends in the Evolution of the Euglenid Pellicle. Evolution 55 (11): 2215-2235. También presente aquí.
- Leander BS y col. 2007. Macroevolution of complex cytoskeletal systems in euglenids. BioEssays 29 (10): 987-1000. Artículo también completo aquí.
- Linton EW & Triemer RE. 1999. Reconstruction of the feeding apparatus in Ploetia costata (Euglenophyta) and its relationships to other euglenoid feeding apparatuses. Journal of Phycology 35 (2): 313-324.
- Simpson AGB y col. 2006. The evolution and diversity of kinetoplastid flagellates. Trends in Parasitology 22 (4): 168-174. Artículo también disponible aquí.
- Yamaguchi A y col. 2012. Morphostasis in a novel eukaryote illuminates the evolutionary transition from phagotrophy to phototrophy: description of Rapaza viridis n. gen. et sp. (Euglenozoa, Euglenida). BMC Evolutionary Biology 12:29 doi:10.1186/1471-2148-12-29.

Últimos comentarios
| Eduardo Baldu en En EEUU los hospitales privado… | |
| Anónimo en Si la ciudadanía no comprende… | |
| Eduardo Baldu en Si la ciudadanía no comprende… | |
| m64bhkhfyj en Si la ciudadanía no comprende… | |
| Anónimo en La desforestación selectiva pu… | |
| Eduardo Baldu en ¿Los humanos somos el centro d… | |
| ateo666666 en ¿Los humanos somos el centro d… | |
| Anónimo en ¿Los humanos somos el centro d… | |
| ateo666666 en El peligroso engaño de que la… | |
| Anónimo en El peligroso engaño de que la… | |
| Eduardo Baldu en ¿Los humanos somos el centro d… | |
| Anónimo González en ¿Los humanos somos el centro d… | |
| ateo666666 en ¿Los humanos somos el centro d… | |
| luisgonzalesm en ¿Los humanos somos el centro d… | |
| ateo666666 en ¿Los humanos somos el centro d… |
Autores

Todos los textos de La Ciencia y sus Demonios se ofrecen bajo licencia Creative Commons Reconocimiento-NoComercial-SinObraDerivada 3.0 .



ClustrMaps

En línea ahora:
 "La Ciencia y sus Demonios" es miembro de la Red Escéptica
"La Ciencia y sus Demonios" es miembro de la Red Escéptica






Síguenos pulsando aquí

“Lo que puede ser afirmado sin pruebas, puede ser descartado sin pruebas”

¡¡Excelente entrada!!
Me gustaMe gusta
Excelente, de verdad.
Me gustaMe gusta
Pero eso no es una forma intermedia entre depredador y fotosintético. Es un microorganismo tanto depredador como fotosíntético.
Como esta es una entrada más especializada y no creo que entren los creatas de turno, he decidido decirlo yo. 😀
Ahora en serio. me fascinan estos «bichos» que se comen las algas y luego aprovechan los cloroplastos. Me quiere sonar que se conoce alguno más, pero pluricelular tipo «gusano» (si, vocabulario técnico o muy técnico 😀 )¿o me equivoco?
Me gustaMe gusta
Valeeee.
Tirón de orejas para mí, por no leer los artículos relacionados antes de comentar.
Ya me he (habéis) contestado.
Me gustaMe gusta
Muchas gracias a todos ^_^
Pues sí, Herbert, la babosa Elysia chlorotica es un buen ladrón de cloroplastos. Parecido es el platelminto Convoluta, pero este se jama algas enteras y las mantiene vivas en su interior, no tiene tanto mérito como hacer funcionar el cloroplasto pero bueh…
En cuanto al creacionista típico, pues sí, es bastante curioso. En entradas generales, sobre todo dedicadas a la parte teórica de la evolución, pues son relativamente habituales. Pero cuando se trata de ejemplos concretos no aparecen, 😀 Por eso me gusta combinar teoría y aplicación práctica; de hecho suelo montar los artículos buscando primero el ejemplo y luego la explicación teórica que sostiene dicho ejemplo 🙄
Me gustaMe gusta
Pues sigue así.
Con ese planteamiento consigues sorprender primero e instruir después.
Que los demás autores no se ofendan, todos tienen un gran estilol
Me gustaMe gusta
Muy bueno, felicitaciones!!
Me gustaMe gusta
estupendo articulo.
Me gustaMe gusta
Cnidus: aquí un tal emilioG y otros no se atreven contigo…. jajajaaj
Me gustaMe gusta
Gracias de nuevo ^_^
Me gustaMe gusta
por fin, estuve buscando por mil años como se alimentaba el flagelado, y por fin lo encontré, y muy buena la informacion
Me gustaMe gusta
Gracias, use mucho de las imágenes y contenido en mis clases.
Me gustaMe gusta